
Морфометрическая и молекулярно-генетическая дифференциация линий и семей медоносной пчелы Apis mellifera caucasica L., разводимых в районе Большого Сочи
 Создание специализированных линий — один из приемов генетического совершенствования и сохранения биоразнообразия пород и популяций медоносной пчелы. Серая горная кавказская порода (Apis mellifera caucasica L.) выделяется среди прочих по комплексу хозяйственно полезных признаков (самый длинный хоботок, высочайшая работоспособность и продуктивность, использование широкого видового состава медоносов, чрезвычайно слабая ройливость, миролюбие и др.). Целью настоящего исследования была сравнительная оценка разнообразия и степени дифференциации линий A m. caucasica на основании данных морфометрического анализа и микросателлитов (МС).
Создание специализированных линий — один из приемов генетического совершенствования и сохранения биоразнообразия пород и популяций медоносной пчелы. Серая горная кавказская порода (Apis mellifera caucasica L.) выделяется среди прочих по комплексу хозяйственно полезных признаков (самый длинный хоботок, высочайшая работоспособность и продуктивность, использование широкого видового состава медоносов, чрезвычайно слабая ройливость, миролюбие и др.). Целью настоящего исследования была сравнительная оценка разнообразия и степени дифференциации линий A m. caucasica на основании данных морфометрического анализа и микросателлитов (МС).
Материалом для исследований служили рабочие пчелы пяти линий (I — линия № 19, с. Медовеевка; II — линия № 13, ПГТ Красная Поляна; III — линия № 12, ПГТ Красная Поляна; IV — линия № 49, с. Калиновое Озеро; V — линия № 34, с. Аибга; n = 728) с пяти пасек в районе Большого Сочи (Краснодарский край). Для морфометрического анализа измеряли длину хоботка (ДХ, мм), ширину 3-го тергита (ШТ, мм) и рассчитывали кубитальный индекс (КИ). Молекулярно-генетические исследования проводили по семи локусам МС (A024, A88, A113, AP043, HB-C16-05, HB-THE-03, HB-C16-01). Степень межсемейной изменчивости по морфометрическим показателям определяли методом двухфакторного иерархического дисперсионного анализа. Генетические различия между семьями по МС оценивали по показателям Fst при парном сравнении. Полученную матрицу использовали для PCA-анализа (principal component analysis). Для количественной оценки межсемейной изменчивости внутри линий рассчитывали индексы Fst, Rst (AMOVA). Степень дифференциации линий по морфометрическим признакам определяли посредством расчета евклидовых дистанций. Полученные значения использовали для построения дендрограммы подобия методом одиночной связи (single linkage) в алгоритме иерархической кластеризации. Степень дифференциации линий по МС оценивали на основании расчета значений генетических дистанций по М. Nei. Дендрограмму подобия строили с помощью метода UPGMA. Для расчетов использовали программное обеспечение STATISTICA, GenAlEx (v. 6.5.1), PAST (v. 3.03). Результаты морфометрического анализа показали наличие достоверных различий между линиями по ДХ и Ш3Т, тогда как в отношении КИ различий не обнаружили. Установлена большая гетерогенность в отношении изучаемых признаков в линиях II и V и, напротив, большая консолидация в линиях III и IV. Пчелы линии I существенно отличались от остальных по обоим показателям, однако характеризовались значительными межсемейными различиями по ДХ. Анализ МС профилей выявил сходные тенденции при оценке степени внутрисемейной и межсемейной изменчивости. В линии I наблюдается избыток гетерозигот (Fjj = -0,048), что можно рассматривать как указание на высокую гетерогенность. Пчелы этой линии характеризовались минимальной индивидуальной (F,t = 0,052) и максимальной межсемейной (Fst = 0,124) изменчивостью. В линиях II-V наблюдали дефицит гетерозигот (Fjj = 0,062-0,128), относительно высокую индивидуальную изменчивость (Fit = 0,143-0,189) и более низкие по отно шению к линии I значения межсемейной изменчивости (Fst = 0,095-0,104). Наименьшие межсемейные различия были отмечены в линиях III и IV (Fst 0,096 и 0,095). Анализ дифференциации изучаемых линий по морфометрическим признакам и МС выявил различия в структуре генеалогического дерева. Дендрограмма, построенная по МС, была отражением географической удаленности изучаемых линий. Структура генеалогического дерева, построенного по морфометрическим признакам, не отражала ни географическую общность (различия) происхождения, ни сходство (различия) в хозяйственно полезных признаках у изучаемых линий. Таким образом, при исследовании морфометрических показателей и МС обнаружены сходные тенденции в оценке внутрисемейной и внутрилинейной изменчивости, однако наблюдались различия при дифференциации линий с использованием двух методов. В дальнейшем комплексный подход позволит с высокой точностью проводить определение не только породы пчел, но и более мелких таксономических групп. Полученные результаты могут быть использованы в селекционной работе по восстановлению чистоты пород медоносных пчел.
Сохранение многообразия пород медоносной пчелы (Apis mellifera L., 1758), а также более мелких таксономических единиц (экотипов и линий) требует глубоких фундаментальных популяционно-генетических исследований состояния их генофонда (1). Внутри вида A. mellifera выделяют около 30 подвидов (2), для обозначения которых используют также термин раса, а в русскоязычной литературе — порода. На территории Российской Федерации разводится несколько пород: среднерусская, краинская (или карника), карпатская (некоторые авторы классифицируют ее как подвид карники 2-го порядка), серая горная кавказская. Последняя по комплексу хозяйственно полезных признаков (самый длинный хоботок, высочайшая работоспособность и продуктивность, использование широкого видового состава медоносов, чрезвычайно слабая ройливость, миролюбие и др.) считается одной из лучших (3). Структура серой горной кавказской породы неоднородна и состоит из популяций, достоверно различающихся по биологическим и хозяйственным признакам (4). Селекционная работа с породой, которая в настоящее время ведется на базе Краснополянской опытной станции (район Большого Сочи), направлена на сохранение генофонда и создание новых специализированных линий с улучшенными хозяйственно полезными качествами (5, 6).
Определение породной принадлежности пчел и их отнесение к той или иной линии — непростая процедура, поскольку различия между линиями в пределах одной породы, как правило, невелики. Кроме того, имеет место спонтанная гибридизация. Преимущественно используется оценка по экстерьерным, физиологическим, этологическим признакам и показателям хозяйственной ценности (2, 7). Наиболее часто измеряют длину хоботка, ширину 3-го тергита и вычисляют кубитальный индекс (8). Была показана возможность дифференциации подвидов медоносной пчелы A. m. mellifera и A. m. carnica с использованием морфометрических методов (9). Однако попытка внутривидовой дифференциации популяций пчел A. m. meda, обитающих в различных географических регионах (на расстоянии 500 км), на основании морфометрических данных оказалась неуспешной (10). Поскольку морфометрические признаки зависят от географического размещения пчел, их возраста, питания и широко варьируют даже в пределах одной породы или популяции, возникает необходимость в разработке подходов, основанных на анализе генотипа.
В последние годы все большую популярность для дифференциации пород, экотипов и линий медоносной пчелы находят молекулярно-генетические методы, включающие исследование аллозимов (11), митохондриальной ДНК (12-14), полиморфизма единичных нуклеотидов (15, 16) и микросателлитов (17-20). Оценка по ДНК-маркерам рассматривается в качестве дополнения к традиционному анализу по морфометрическим критериям (21, 22). Ранние работы, в которых ДНК-анализ использовался для идентификации внутривидовых таксономических единиц медоносной пчелы, базировались на определении уникальных (приватных) для соответствующей линии аллелей (23). Затем применение байесовских методов сделало возможным генетическую кластеризацию индивидуумов (подвидов и более мелких таксонов) на основе генотипа по множеству несцепленных локусов (24). Метод высокоэффективен не только при определении подвидов, относящихся к различным эволюционным ветвям A. mellifera (25, 26), но и для установления происхождения более близких таксонов (27).
В представляемой работе мы показали, что структура генеалогического дерева, построенного по морфометрическим признакам, не отражала ни географическую общность происхождения, ни сходство в хозяйственно полезных признаках у изученных линий. Следовательно, для их адекватной характеристики и дифференциации необходимо сочетание морфометрических и молекулярно-генетических методов. Использованный нами комплексный подход позволит с высокой точностью определять не только породы пчел, но и более мелкие таксономические группы. Полученные результаты могут быть использованы в работе по восстановлению чистоты пород медоносных пчел.
Целью настоящей работы стала сравнительная оценка разнообразия и степени дифференциации линий Apis mellifera caucasica на основании данных морфометрического анализа и микросателлитов.
Методика. Исследования проводили в 2014 году. В качестве материала использовали рабочих пчел серой горной кавказской породы различных линий, разводимых на пяти пасеках в районе Большого Сочи (Краснодарский край): I — линия № 19 (с. Медовеевка), II — линия № 13 (ПГТ Красная Поляна), III — линия № 12 (ПГТ Красная Поляна), IV — линия № 49 (с. Калиновое Озеро) и V — линия № 34 (с. Аибга). Для анализа отбирали 5 семей каждой линии по 22-30 пчел в семье (25 семей, n = 728), в том числе I — n = 149, II — n = 147, III — n = 147, IV — n = 137, V — n = 148. Пчел консервировали в 96 % спирте и хранили при +4 °С.
Измеряли длину хоботка (ДХ, мм), ширину 3-го тергита (ШТ, мм) и рассчитывали кубитальный индекс (КИ) как отношение между большей и меньшей сторонами третьей кубитальной ячейки переднего крыла (8).
ДНК выделяли с помощью экстракции перхлоратом (28), материалом служила голова рабочей пчелы. Семь локусов микросателлитов (МС) — A024, A88, A113, AP043, HB-C16-05, HB-THE-03, HB-C16-01 амплифици-ровали в одной мультиплексной реакции с использованием флуоресцентно меченных праймеров. Реакции проводили в конечном объеме 15 мкл. В пробирки вносили по 14 мкл реакционной смеси (1 мкл 10* ПЦР-буфера, 0,5 мкл 2 мМ раствора dNTPs, 0,7 мкл 100 мМ раствора DMSO, 4,6 мкл 10 мМ смеси праймеров, 0,1 мкл Таq-полимеразы, 7,1 мкл бидистиллированной воды) и добавляли 1 мкл (50-100 нг) исследуемой геномной ДНК. Состав ПЦР-буфера: 16,6 мМ (NH4)2SO4, 67,7 мМ Трис-НС1 (рН = 8,8), 0,1 объема Tween 20. ПЦР выполняли в следующем режиме: начальная денатурация 8 мин при 94 °С; отжиг 2 мин при 55 °С; полимеризация 2 мин при 72 °С; 30 циклов — денатурация 30 с 94 °С, отжиг 35 с при 55 °С, элонгация 40 с при 72 °С; элонгация 20 мин при 72 °С. Разделение продуктов ПЦР выполняли на генетическом анализаторе ABI3130xl («Applied Biosystems», США) с использованием внутреннего стандарта ROX. Элек-трофореграммы обрабатывали с помощью программного обеспечения Ge-neScan v. 3.7 и Genotyper v. 3.7 («Applied Biosystems», США).
Межсемейную изменчивость по морфометрическим показателям оценивали методом двухфакторного иерархического дисперсионного анализа с помощью программного обеспечения Statistica v. 6 (29). В качестве фиксированных эффектов использовали принадлежность к линии и семье (иерархический подчиненный фактор). Рассчитывали Mst:
![]()
где F — дисперсионное отношение, n* — среднее геометрическое объема выборок. Величина Mst может принимать значения от 0 до 1 и оценивать меру морфологической дифференциации между отдельными популяциями и, по сути, представляет собой аналог Fst (30).
Генетические различия между семьями медоносной пчелы в отношении МС оценивали по показателю Fst (31) при парном сравнении и на основании генетических дистанций по M. Nei (32). Полученные попарные матрицы использовали для построения распределения центроидов семей в пространстве первых двух главных координат (PCA, principal component analysis). Для количественной оценки степени межсемейной изменчивости внутри линий рассчитывали идексы Fst, Rst (AMOVA) (33), используя программное обеспечение GenAlEx v. 6.5 (34).
Степень дифференциации пяти изучаемых линий A m. caucasica по морфометрическим признакам оценивали посредством расчета евклидовых дистанций. Полученные значения использовали для построения дендрограммы подобия методом одиночной связи (single linkage) в алгоритме иерархической кластеризации (29). Оценку степени дифференциации линий по МС проводили на основании расчета значений генетических дистанций по M. Nei (32). Дендрограмму подобия строили методом UPGMA при помощи программного обеспечения PAST (v. 3.03). Дендрограмму проверяли на устойчивость топологии кластеров посредством проведения 999 перму-таций bootstrap-методом.
Результаты. При формировании каждая из изученных линий пчел была строго ориентирована на получение материала с заданными хозяйственно полезными признаками. Матки линии III обладают высокой яйценоскостью в период подготовки к основному медосбору, а семьи — повышенной продуктивностью по меду. Линии IV и V характеризуются ранним развитием по сравнению с линиями I, II и III. Пчелы линии IV менее зимостойки, в то время как линия II выделяется повышенной, а линия I — высокой зимостойкостью.
Были выявлены достоверные различия между линиями по двум из трех оцененных признаков — длине хоботка и ширине 3-го тергита, тогда как по значениям кубитального индекса различий между линиями мы не обнаружили (табл. 1). У пчел линии I по сравнению с другими изучаемыми линиями величина ДХ оказалась на 0,05-0,07 мм меньше (p < 0,001), а Ш3Т — на 0,04-0,11 мм больше (p < 0,001). Максимальную длину хоботка отмечали у пчел линии III. У пчел линий III и IV Ш3Т была на 0,08-0,11 мм меньше, чем у других линий (p < 0,001). Обнаруженные различия могут быть следствием разной стратегии при селекционно-племенной работе с этими линиями.
1. Морфометрические показатели у различных линий медоносных пчел серой горной кавказской породы (Apis mellifera caucasica L.) (X+x, Краснодарский край, 2014)

Примечание. ДХ — длина хоботка, Ш3Т — ширина 3-го тергита, КИ — кубитальный индекс. Описание линий см. в разделе «Методика».
а, ь, c, d, e, f, g, h p < 0,001, i p < 0,01 (достоверность различий между значениями, обозначенным одной и той же буквой).
Результаты двухфакторного иерархического дисперсионного анализа подтвердили наличие достоверных различий по признакам ДХ и Ш3Т как между линиями, так и между семьями в пределах каждой из линий.
Формирование кластеров по линейному принципу наблюдалось для трех из пяти исследованных линий (рис. 1). Только семьи линии I распределились внутри одного обособленного 1-го кластера, однако они характеризовались наибольшей изменчивостью по показателю ДХ (Сдх = 1,11 %) по сравнению с другими линиями (Сдх = 0,69-0,86 %) при средней гетерогенности по Ш3Т (СкщэТ = 2,49 %). Семьи линий III и IV формировали два консолидированных кластера (соответственно Сдх 0,74 и 0,69 % и Сшзт 2,15 и 2,33 %), которые частично перекрывались между собой. У пчел линий II и V сильно варьировали оба признака (Сдх — 0,79 и 0,86 %, Сшзт — 2,66 и 2,52 %). Достоверные различия (р < 0,05) между семьями внутри линий наблюдались по признаку ДХ в линии II (Mst = 0,1359) и признаку Ш3Т в линиях II (Mst = 0,2869) и V (Mst = 0,1525). Межлинейные различия были достоверны (р < 0,05) для показателей ДХ (Mst = 0,1453) и Ш3Т (Mst = 0,1097), в то время как по КИ достоверных различий мы не выявили.
В целом следует отметить большую гетерогенность в отношении изучаемых признаков у пчел линий II и V и, напротив, большую консолидацию у пчел линий III и IV. Пчелы линии I существенно отличались от остальных по обоим показателям, однако характеризовались значительными межсемейными различиями по ДХ.
Анализ МС профилей выявил сходные тенденции в определении степени внутрисемейной и межсемейной изменчивости (табл. 2).
В линии I (единственной из исследованных) наблюдался избыток гетерозигот (отрицательное значение Fis), что можно рассматривать как
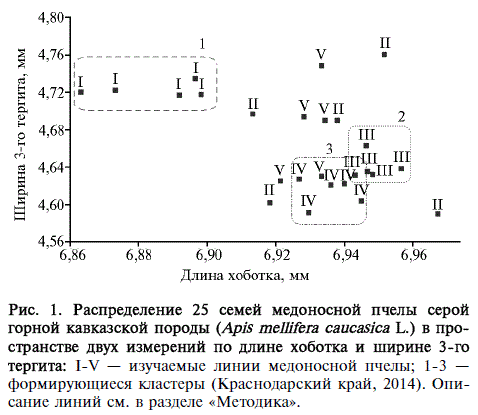
Рис. 1. Распределение 25 семей медоносной пчелы серой горной кавказской породы (Apis meUifsra caucasica L.) в пространстве двух измерений по длине хоботка и ширине 3-го тергита: I-V — изучаемые линии медоносной пчелы; 1-3 — формирующиеся кластеры (Краснодарский край, 2014). Описание линий см. в разделе «Методика».
2. Степень внутрисемейной и межсемейной (внутри линий) генетической изменчивости у медоносной пчелы серой горной кавказской породы (Apis mellifera caucasica L.) по семи локусам микросателлитов (Краснодарский край, 2014)
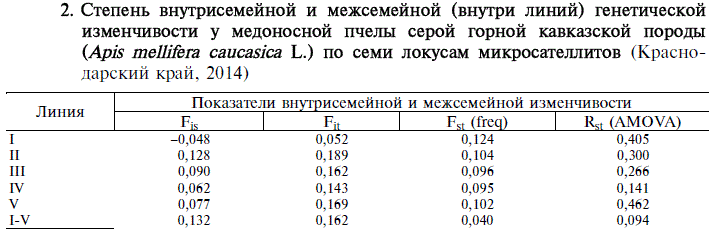
Примечание. Расчеты проводили внутри каждой из линий с использованием семьи в качестве субпопуляции, а также для выборки в целом (I-V) с использованием линии в качестве субпопуляции. Описание линий и локусов микросателлитов см. в разделе «Методика».
указание на высокую гетерогенность. Пчелы этой линии характеризовались минимальной индивидуальной изменчивостью по отношению к выборке в целом (значение Fit было в 2,75-3,63 раза ниже по сравнению с линиями II-V) и максимальной межсемейной изменчивостью. У пчел линии II-V имелся дефицит гетерозигот (положительные значения индекса Fis), отмечалась относительно высокая индивидуальная изменчивость по сравнению с выборкой в целом и более низкая по отношению к линии I межсемейная изменчивость. Наименьшие межсемейные различия, оцененные по показателям Fst и R^ (AMOVA), выявили в линиях III и IV.
Результаты PCA-анализа показали наибольшее сходство семей внутри линии IV, в то время как линии I и II характеризуются наибольшими межсемейными различиями (рис. 2).
Анализ дифференциации изучаемых линий выявил различия в структуре генеалогического дерева, построенного для семи локусов МС и трех морфометрических признаков (рис. 3). Дендрограмма, построенная по МС, отражала географическую удаленность изучаемых линий (см. рис. 3, А). Формирование двух главных кластеров могло быть следствием географической изолированности линий I, II и III, расположенных в окрестностях Красной Поляны (первая пасека находится на южном склоне хребта Ачишхо высотой 2391 м, вторая и третья расположены в ущелье реки Мзымта), от линии IV (пасека расположена на расстоянии 15-25 км от трех перечисленных и отделена от них хребтом Алек высотой 1000 м) и линии V (находится в верховьях реки Псоу, удалена от первой, второй и третьей пасек на 10-16 км и отделена от них хребтом Аибга высотой 2460 м). Существенные генетически различия между линиями IV и V могли быть обусловлены их географической изолированностью. Пасеки, на которых разводят эти линии, расположены на расстоянии около 30 км друг от друга и отделены хребтом Алек, который протянулся на 20 км между реками Мзымта и Сочи на расстоянии 15 км от берега Черного моря, и хребтом Аибга, протянувшимся с северо-запада на юго-восток на 23 км и разделяющим среднюю часть долины Мзымты и верховья Псоу.
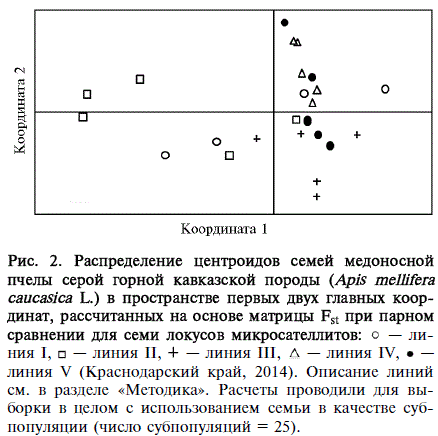
Рис. 2. Распределение центроидов семей медоносной пчелы серой горной кавказской породы (Apis mellifeia caucasica L.) в пространстве первых двух главных координат, рассчитанных на основе матрицы Fst при парном сравнении для семи локусов микросателлитов: о — линия I, □ — ЛИНИЯ II, + — ЛИНИЯ III, А — линия IV, • — линия V (Краснодарский край, 2014). Описание линий см. в разделе «Методика». Расчеты проводили для выборки в целом с использованием семьи в качестве субпопуляции (число субпопуляций = 25).
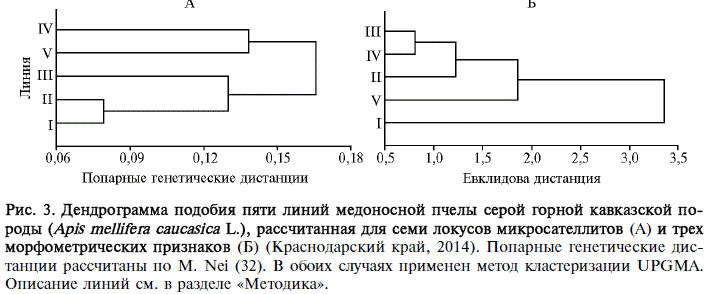
Рис. 3. Дендрограмма подобия пяти линий медоносной пчелы серой горной кавказской породы (Apis mellifera caucasica L.), рассчитанная для семи локусов микросателлитов (А) и трех морфометрических признаков (Б) (Краснодарский край, 2014). Попарные генетические дистанции рассчитаны по M. Nei (32). В обоих случаях применен метод кластеризации UPGMA. Описание линий см. в разделе «Методика».
Структура генеалогического дерева, построенного по морфометрическим признакам, не отражала ни географическую общность (различия) происхождения, ни сходство (различия) в хозяйственно полезных признаках изучаемых линий.
Таким образом, нами обнаружены сходные тенденции при оценке внутрисемейной и внутрилинейной (между семьями) изменчивости на основе морфометрических показателей и микросателлитов. Однако данные о степени дифференциации и характере генеалогических связей между линиями, полученные с использованием двух систем анализа, существенно различались. Возможно, подобные расхождения обусловлены тем, что большинство различий по традиционным экстерьерным признакам (морфологические промеры) укладываются в пределы сезонных колебаний и малопригодны для идентификации локальных популяции. Кроме того, они в большей степени подвержены давлению отбора факторов внешней среды, не связанных с воздействием направленного искусственного отбора. Микросателлиты в большинстве случаев селекционно нейтральны и отражают более отдаленные эволюционные события. Следовательно, для получения наиболее полной информации о состоянии и динамике популяций необходимо учитывать результаты, полученные с использованием обоих методов.
М.С. ФОРНАРА, А.С. КРАМАРЕНКО, С.В. СВИСТУНОВ, Е.М. ЛЮБИМОВ,
С.С. СОКОЛЬСКИЙ, Н.А. ЗИНОВЬЕВА






